






Les poissons des récifs coralliens
Extrait des "Lettres récifales" nr 6 - juillet 1997
 |
| Les récifs coralliens abritent une faune foisonnate et variée. Les poissons, particulièrement nombreux et colorés, représentent un enjeu économique important pour les pays tropicaux. Photo : Marcel Staebler |
Saviez-vous que quelques kilomètres de récifs coralliens
abritent plus d'espèces de poissons que toute la Méditerranée
? L'économie côtière des pays tropicaux est en grande
partie fondée sur cette richesse. Faire fructifier un tel trésor
exige de comprendre ce qui conditionne l'abondance des poissons.
Aussi les écologues s'efforcent-ils de décrypter les étapes
du cycle de vie des poissons coralliens qui les mènent d'un voyage
erratique dans l'océan à la reproduction dans les récifs.
|
LES CORAUX DES R?CIFS Ce sont des animaux relativement primitifs appartenant au groupe des madr?poraires. Ils vivent en solitaires ou en colonies. Chaque individu, le polype, comprend une cavit? gastrique qui se prolonge par un pharynx et une bouche entour?e de tentacules. Ceux-ci servent ? capturer les proies et les d?bris organiques dont les coraux se nourrissent. La symbiose avec des algues photosynth?tiques assure un compl?ment nutritif. Dans une colonie, chaque polype est en relation avec ses voisins par un tissu mou, le ccenosarque. Les polypes s?cr?tent autour d'eux un squelette calcaire qui finit par constituer le r?cif au fur et ? mesure de la croissance et de la mort des coraux. |
Les r?cifs coralliens sont les plus grandes constructions jamais ?labor?es
par des ?tres vivants : les coraux madr?poraires (voir l'encadr? ? Les
coraux des r?cifs ?). Ces animaux marins h?bergent dans leurs tissus des
algues unicellulaires, les zooxanthelles, qui vivent en symbiose avec eux.
Les avantages sont mutuels . le corail constitue un abri pour l'algue, tandis
que celle-ci fournit de la mati?re organique qui contribue ? l'alimentation
de l'h?te. Cette association pr?sente un inconv?nient. Pour effectuer la
photosynth?se, les zooxanthelles ont besoin d'?nergie lumineuse. Aussi les
coraux ne peuvent-ils vivre que dans les eaux superficielles de l'oc?an o?
la lumi?re p?n?tre. Ils exigent une temp?rature minimale de l'eau de 20
°C, leur croissance ?tant optimale entre 23 et 27 °C. Les
?cosyst?mes que forment les r?cifs coralliens, v?ritables ? oasis d'abondance
dans un d?sert d'oc?ans ?, sont beaucoup plus productifs que l'oc?an environnant.
Mesur? en taux de carbone fix? par photosynth?se, leur rendement est d'environ
dix grammes par m?tre carr? et par jour, soit l'?quivalent de la productivit?
moyenne d'un champ cultiv?. Comme les for?ts tropicales auxquelles ils sont
souvent compar?s les r?cifs coralliens constituent une source de nourriture
et un abri pour un nombre consid?rable d'esp?ces animales: un r?cif de
quelques kilom?tres peut renfermer quatre cents esp?ces de poissons, soit
autant que toute la M?diterran?e et compte de surcro?t des dizaines d'esp?ces
de coraux, de mollusques (porcelaines, c?nes, t?r?bres, etc.), d'?chinodermes
(oursins, synaptes, ?toiles de mer) ou de crustac?s (crabes et crevettes).
L'enjeu ?conomique et social des r?cifs coralliens est d'ailleurs directement
li? ? la richesse de leur faune. La p?che, souvent artisanale, reste l'activit?
essentielle de nombreuses r?gions c?ti?res ? r?cifs coralliens - soit
15 % des syst?mes marins c?tiers, ou 617 000 km2 , en particulier
dans le Pacifique (Polyn?sie) et en Asie du Sud-Est (Philippines, Indon?sie)
Poissons, crustac?s, mollusques, etc., sont destin?s ? l'alimentation,
? l'artisanat et ? l'aquariophilie. Une exploitation humaine grandissante
des r?cifs, ajout?e aux causes naturelles de destruction (cyclones, maladies
telle que la ? mort blanche ? des coraux, pr?dation des coraux par l'?toile
de mer Acanthaster plancii fait que de plus en plus de r?cifs coralliens
connaissent des d?gradations irr?versibles.
Les r?cifs coralliens se caract?risent surtout par une faune de poissons
d'une tr?s grande diversit?. Environ quatre mille esp?ces y ont ?t? recens?es,
soit 30 % des poissons marins et de nouvelles esp?ces sont encore d?crites
r?guli?rement. Ces poissons, dont la beaut?, la richesse des couleurs,
l'extravagance des formes et la diversit? des comportements sont incomparables,
vivent pr?s du substrat r?cifal. La transparence de l'eau et la proximit?
du fond, souvent couvert d'invert?br?s color?s, expliquent peut-?tre ces
couleurs vives : elles les camouflent par mim?tisme, ou facilitent la reconnaissance
ou l'attraction des cong?n?res.
Le trait probablement le plus m?connu des poissons coralliens, et pourtant
le plus d?terminant pour la biodiversit? des r?cifs, concerne le d?but
de leur vie. Plus de 99 % des esp?ces - comme beaucoup d'autres poissons
marins, sauf les requins et les raies - passent par deux phases fondamentalement
diff?rentes : tout d'abord, les larves voire les oeufs tout juste f?cond?s,
ont une vie oc?anique et planctonique (ils sont dispers?s par les courants
marins) ; puis les juv?niles et les adultes nagent pr?s du r?cif. La premi?re
p?riode influence aussi bien la r?partition des esp?ces ? l'?chelle mondiale
que leur devenir dans les r?cifs. De m?me, le changement radical de mode
de vie entre la phase larvaire et la phase adulte est une ?tape d?cisive
pour le peuplement des r?cifs : le ? recrutement ?.
L'un des facteurs qui a contribu? ? la r?partition g?ographique actuelle
des poissons coralliens est leur possibilit? de dispersion lors de la phase
larvaire oc?anique. Il existe pour les r?cifs deux grandes provinces biog?ographiques,
la province atlantique, avec notamment les Cara?bes, et la province indo-pacifique,
vingt fois plus ?tendue, du golfe de Suez jusqu'au Pacifique Est. Leur origine
tient ? la s?paration des deux bassins oc?aniques du Pacifique et de l'Atlantique
voil? deux ? cinq millions d'ann?es. Dans l'Indo-Pacifique, la plus riche
en esp?ces, le nombre maximal de poissons se situe dans l'archipel des Philippines
(deux mille esp?ces recens?es). Ce nombre d?croit ensuite selon la longitude
et la latitude. Mais il reste cependant plus grand le long des continents
(Australasie et Afrique) qu'au niveau des ?les, car les grands espaces oc?aniques
repr?sentent une certaine barri?re ? la dispersion des esp?ces qui, comme
les poissons des r?cifs, vivent pr?s du fond. Seulement huit cents esp?ces
de poissons existent en Polyn?sie fran?aise alors que Madagascar, par exemple,
en compte mille deux cents. Inversement, l'isolement g?ographique favorisant
la naissance de nouvelles esp?ces, l'end?misme (la quantit? d'esp?ces
restreintes ? une r?gion) est plus important dans les ?les ?loign?es
des continents.
Comment se d?roule la reproduction sexu?e d'o? proviennent les larves ?
Des familles distinctes de poissons ont un comportement reproducteur similaire(3).
Certaines d'entre elles, comme les poissons-chirurgiens ou les muges, se reproduisent
en formant des bancs importants dans les passes et les fa?ades oc?aniques
des r?cifs, endroits les plus expos?s ? l'influence oc?anique. Les cellules
sexuelles m?les et femelles (gam?tes) sont ?mises dans l'eau, o? s'effectue
ensuite la f?condation.
Chez d'autres esp?ces, tels les poissons demoiselles et les poissons-papillons,
des couples font de br?ves ascensions vers la surface et y lib?rent leurs
gam?tes. Chez un tiers des esp?ces, les femelles pondent leurs oeufs non
f?cond?s sur un substrat dur, comme certains poissons demoiselles, ou dans
un nid grossier creus? dans le s?diment et quelles prot?gent vigoureusement,
tels les poissons balistes. N?anmoins, quel que soit le lieu de f?condation,
benthique ou p?lagique, les larves sont p?lagiques et ne retournent dans
le r?cif, pour le recrutement qu'? l'issue d'un s?jour de deux semaines
? plus de trois mois en plein oc?an.
Des milliers, voire des millions d'oeufs de moins d'un millim?tre de diam?tre
produits par la f?condation, naissent des larves minuscules qui sont emport?es
dans l'oc?an par les courants. Elles se nourrissent d'abord gr?ce ? leur
sac vitellin, riche en r?serves nutritives, puis capturent de petites proies
planctoniques.
A ce stade, les larves sont transparentes et se d?placent par saccades en
battant rapidement de leur queue fine comme un flagelle (? leur ?chelle
l'eau est relativement visqueuse). Les jeunes larves ne peuvent donc r?sister
aux mouvements oc?aniques ef font partie du plancton oc?anique. Certaines
larves s'adaptent ? la vie planctonique en d?veloppant une morphologie,
une physiologie ou des comportements appropri?s (fig. 1). A la fin de leur
s?jour dans apparaissent sur le corps ? des endroits propres ? chaque esp?ce,
ce qui permet son identification.
|
|
|
Fig. 1: Lors de la reproduction, les femelles des poissons coralliens pondent des oeufs qui donnent naissance ? de minuscules larves. Les poissons coralliens d?butent dans la vie par une phase larvaire planctonique de quelques semaines de qui se d?roule en plein oc?an. Au terme e cette phase, les individus d'une m?me esp?ce peuvent se retrouver disperser en diff?rentes zones de l'oc?an. On voit ci-dessus quelques exemples de larves. Des modif ications telles que les ?pidenticul?es des nageoires dorsales et pelviennes de certaines larves facilitent le transport par les courants et constituent vraisemblablement une adaptation ? la vie planctonique. (Clich? des auteurs) |

Les larves sont alors pr?tes pour le grand bouleversement de leur cycle
de vie : leur recrutement parmi les poissons du r?cif et leur m?tamorphose
en adultes. Mais toutes n'arrivent pas ? destination : pr?dation, le manque
de nourriture, l'entra?nement par certains courants dans des zones oc?aniques
d?favorables provoquent une h?catombe. Sur les dizaines de milliers d'oeufs
que pond chaque femelle, il ne subsiste que quelques centaines de larves ?g?es
ph?nom?ne d'ailleurs quasi g?n?ral chez les poissons et beaucoup d'invert?br?s
marins. Pour celles qui restent, la transition entre la vie dans l'oc?an
et la vie sur le r?cif se d?roule par le biais d'une m?tamorphose. Les
larves de poissons r?cifaux acqui?rent progressivement toutes les aptitudes
physiologiques pour se transformer en juv?niles color?s. La m?tamorphose
s'op?re g?n?ralement au contact des r?cifs, peut-?tre sous l'effet de
stimuli sensoriels particuliers. Mais elle peut ?galement se produire au
large. Il est donc essentiel que les larves soient suffisamment proches d'un
r?cif pour, venir le coloniser et y poursuivre leur cycle de vie. Celles
qui en restent ?loign?es risquent de se m?tamorphoser en plein oc?an.
Leur nouvelle coloration les rend alors extr?mement vuln?rables vis-?-vis
des nombreux pr?dateurs p?lagiques.
N?anmoins, il semble que les larves parviennent ? ?chapper ? une mort
pr?matur?e en retardant leur m?tamorphose. Des travaux r?alis?s en 1992
par Benjamin Victor aux Cara?bes ont montr?, chez une esp?ce de girelle
du genre Thalassoma, que la dur?e de vie larvaire peut varier selon
les individus. Certains chercheurs, comme Jeff Leis, de l'Australian Museum,
pensent que les larves pourraient ralentir leur d?veloppement quand elles
se trouvent au large et augmenter ainsi leurs chances de rencontrer un r?cif.
Compte tenu de la forte mortalit? larvaire provoqu?e par la vie au large
et le retour au r?cif, on peut se demander quel est, pour les esp?ces de
poissons, l'int?r?t de la phase p?lagique ? Pourquoi l'?volution n'a-t-elle
pas plut?t s?lectionn? un cycle de vie enti?rement r?cifal ? Un premier
courant d'id?es invoque les avantages adaptatifs de la dispersion des oeufs
et des larves, une diaspora oc?anique qui permet de diminuer les risques
de mortalit? associ?s ? des perturbations locales subies par un r?cif(4).
Ce m?canisme augmente ?galement l'aire de r?partition g?ographique de
l'esp?ce(s). Une th?se voisine propos?e en 1981 par Georges Barlow, de
l'universit? de Californie, sugg?re que la naissance et la disparition de
chaque r?cif corallien ont favoris? la s?lection d'une phase de dispersion
larvaire, car les esp?ces auraient pu, par ce biais, ?viter les risques
d'extinction(6). La dispersion des larves rendrait donc les esp?ces de poissons
coralliens moins sensibles ? la disparition des r?cifs. Pour les sp?cialistes
de l'?volution, cette hypoth?se peut ?tre associ?e au mod?le de m?tapopulation
utilis? en ?cologie : une m?tapopulation est soit une population fragment?e,
soit un ensemble de populations qui ?voluent conjointement du fait d'?changes
g?n?tiques et de migrations. Une dispersion importante permet de contrebalancer
des extinctions localis?es. Un second courant d'id?es explique la dispersion
des larves de poissons en milieu p?lagique en consid?rant le milieu r?cifal
comme un environnement d?favorable ? leur survie. Cet argument a ?t? expos?
en 1978 par Bob Johannes(7), du CSIRO (Commonwealth Scientific and Industrial
Research Organization, l'?quivalent du CNRS en Australie). Selon ce biologiste,
les poissons pr?dateurs, qui repr?sentent 65 % ? 70 % des esp?ces, exercent
une forte pression s?lective dans le r?cif : les esp?ces menac?es doivent
s'adapter ou dispara?tre. La phase larvaire oc?anique serait alors une r?ponse
? la pression de la pr?dation. De plus, certaines hypoth?ses sugg?rent
que l'oc?an procure des conditions relativement favorables au d?veloppement
des larves: la nourriture planctonique y est suffisante, la comp?tition r?duite
du fait de l'espace et les pr?dateurs moins nombreux. T?moins du caract?re
adaptatif de cette ?chapp?e vers l'oc?an, certains sites de ponte correspondent
aux emplacements o? les oeufs et les larves ont le plus de chances d'?tre
entra?n?s vers le large par des courants sortant du r?cit Johannes sugg?re
en outre que ces courants sont souvent associ?s ? des tourbillons oc?aniques.
Ceux-ci agiraient comme des attracteurs maintenant les larves ? proximit?
du r?cif jusqu'au moment o?, ayant atteint une taille et des capacit?s
natatoires suffisantes, elles pourraient nager vers le r?cif pour le coloniser
(voir l'encadr?
|
Le voyage des larves : Partir pour mieux revenir Dans certaines situations, les larves de poissons peuvent demeurer ? proximit? du r?cif o? elles sont n?es. Cela rejoint l'hypoth?se de Bob Johannes sugg?rant l'existence de tourbillons oc?aniques jouant le r?le d'? attracteurs ? des larves les maintenant ? proximit? du r?cif. Ainsi, dans le cas des ?les r?cifales, des chercheurs d'Hawa? comme George Boelhert, de la National Marine Fisheries Service (NOAA), ont analys? la distribution des larves en relation avec la circulation des masses d'eau autour des ?les(14). Ils ont g?n?ralement montr? qu'un courant r?gulier arrivant du large sur une ?le engendre des tourbillons en aval de celle-ci et que les larves de poissons ont tendance ? y ?tre concentr?es. N?anmoins, de tels tourbillons peuvent devenir instables et se d?tacher de l'?le si le courant augmente (voir le sch?ma ci-dessous).
D'autres ?tudes r?alis?es par l'un d'entre nous (P. Doherty) avec
des pi?ges lumineux permettant de capturer les larves ont indiqu?
?galement que celles-ci se regroupent plut?t en aval de l'?le par
rapport au courant. Le m?me processus de r?tention des larves pr?s
des r?cifs peut ?tre observ? pour ceux qui sont situ?s le long des
continents, comme la Grande Barri?re. |
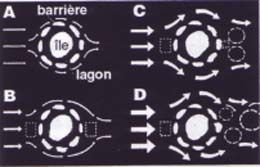
Les p?r?grinations des larves au gr? des courants sont ? l'origine
d'?changes g?n?tiques entre des populations parfois ?loign?es.
Les larves s'adaptent-elles donc ? la pr?dation et aux risques du milieu
r?cifal ? Ou s'agit-il d'une adaptation n?cessaire ? la dispersion et la
survie de l'esp?ce ? Ces deux hypoth?ses ne sont pas contradictoires : en
supposant que les contraintes de la vie dans les r?cifs aient favoris? la
phase oc?anique, les avantages de la dispersion seraient acquis par la m?me
occasion. Les r?les de la dispersion des larves ?tant mieux cern?s, il
reste ? comprendre comment elle modifie, pour chaque esp?ce, la composition
des populations li?es aux diff?rents r?cifs d'une r?gion. Autrement dit,
quels sont les ?changes de g?nes dus ? la reproduction sexu?e entre ces
populations ? Cet aspect est d?terminant pour conna?tre la fa?on dont ces
populations ?voluent et pour am?liorer la gestion des p?ches. Malheureusement,
peu d'?tudes ont ?t? r?alis?es dans ce domaine. Nous avons effectu?
l'une d'entre elles en 1992 sur un poisson-chirtirgien (Acanthurus triostegus)
et un poisson-demoiselle (Dascyllus aruanus). Le cadre en ?tait la
Polyn?sie fran?aise, compos?e de cent dix-huit ?les dont la plupart sont
entour?es de r?cifs coralliens.
 |
| Fig. 2: Les p?rigrinations des larves au gr? des courants marins modifient, pour chaque esp?ce, la composition des populations vivant dans les diff?rents r?cifs. Sur la Grande Barri?re de corail d'Australie, les d?placements et les brassages de g?nes (li?s ? la reproduction sexu?e) entre populations sont d'autant plus intenses que la dur?e de vie larvaire est longue. En effet, comme l'indique ce graphique, ce temps de maturation est corr?l? ? une diminution des diff?rences g?n?tiques entre les populations du Nord et du Sud de la Grande Barri?re (s?par?es par environ 1000 kilom?tres). La dur?e de vie larvaire peut ?tre connue en analysant les stries d'accroissement journalier d?pos?es sur des pi?ces calcifi?es de l'oreille interne, les otolithes. La diff?renciation g?n?tique est ?tablie en analysant certaines enzymes de chaque individu d'une population (plus de 400 individus). A chaque individu est assign?e une sorte de fiche g?n?tique permettant des comparaisons entre populations. |
Pour la premi?re esp?ce, les r?sultats indiquent que les flux g?niques
entre les r?cifs insulaires sont tr?s r?duits (moins de 1 % de la population)
bien que la dur?e de la phase larvaire oc?anique soit relativement longue
(soixante jours) : les populations sont donc g?n?tiquement assez diff?rentes.
En revanche, chez le poisson-demoiselle, dont la phase oc?anique ne dure
que vingt-cinq jours, les populations sont g?n?tiquement uniformes. Ce r?sultat
suppose d'importants brassages pr?sents ou pass?s, li?s ? des successions
d'extinction et de colonisation massive qui ont homog?n?is? les caract?res
g?n?tiques sur l'ensemble des populations de la Polyn?sie. Les flux g?niques
s'effectueraient probablement au gr? des fluctuations des courants oc?aniques
circulant entre les ?les(8-9). Il s'av?re que, dans un syst?me constitu?
de r?cifs ?pars, l'?l?ment primordial qui d?termin? les ?changes de
g?nes entre les populations de poissons n'est pas la dur?e de la phase larvaire
mais les variations des courants reliant les r?cifs. A l'oppos?, dans un
syst?me semi-continu comme la Grande Barri?re d'Australie, o? les r?cifs
se sont align?s sur deux mille kilom?tres le long du plateau continental,
avec des passages plus ou moins larges entre eux, les ?changes de g?nes
entre populations pourraient d?pendre de la dur?e de la phase larvaire oc?anique.
C'est ce que sugg?re une ?tude similaire ? la pr?c?dente, que nous avons
r?alis?e sur sept esp?ces de poissons de la Grande Barri?re : l'augmentation
de la dur?e de vie des larves est corr?l?e ? une diminution des diff?rences
g?n?tiques des populations de poissons; cette dur?e favoriserait donc les
capacit?s migratrices des larves entre les diff?rents r?cifs (10) (fig.
2).
Toutefois, les flux g?niques restent limit?s et ne permettent pas de cr?er
une population unique ? l'?chelle de la Grande Barri?re. La question des
?changes g?niques entre populations est plus cruciale qu'il n'y para?t.
La p?che et la d?gradation des milieux c?tiers entra?nent non seulement
une diminution des stocks de poissons, mais parfois aussi une r?duction de
l'aire de r?partition naturelle des esp?ces (m?me si la p?che reste essentiellement
artisanale dans ces zones). Aussi la connaissance de la phase larvaire oc?anique
appara?t - elle maintenant indispensable pour une gestion rationnelle des
ressources, notamment en milieu corallien. En voici deux exemples.
Faut-il g?rer les poissons coralliens en prot?geant chaque r?cif,
ou se limiter ? quelques sites servant de r?serve ?
Nous avons vu qu'en Polyn?sie fran?aise les populations de certaines esp?ces
sont relativement isol?es les unes des autres. Les quelques migrations sporadiques
sont importantes pour la survie de l'esp?ce, mais elles ne constituent pas
un apport d?mographique important. Pour ces esp?ces, le stock de chaque
?le se renouvelle essentiellement ? partir de son propre peuplement. Dans
un tel cas d'autorecrutement, ces esp?ces devraient donc ?tre g?r?es ?
l'?chelle de chaque ?le afin de permettre le maintien d'un stock de g?niteurs.
A l'oppos?, quand le syst?me est domin? par de nombreux ?changes entre
les r?cifs, comme dans le cas de la Grande Barri?re d'Australie, il est
possible de ne prot?ger qu'un nombre restreint de r?cifs, mais dans leur
totalit?. Des stocks importants de g?niteurs pourraient s'y constituer,
dont les oeufs puis les larves se disperseraient vers les autres r?cifs.
Cette question rejoint le probl?me de la structure spatiale des r?serves
naturelles: faut-il faire une grande r?serve ou plusieurs petites ? Pour
les poissons des r?cifs, il n'y a probablement pas de r?ponse unique etil
convient de r?soudre ce probl?me en tenant compte des caract?ristiques
propres ? chaque milieu. Venons en maintenant au recrutement, deuxi?me phase
d?terminante de la vie des poissons coralliens. Comment les larves retrouvent-elles
les r?cifs ? Existe-t-il un ph?nom?ne de ? homing ? vers les r?cifs
comme pour les saumons remontant les rivi?res ? Ont-elles une capacit? de
rep?rage du champ magn?tique, comme certains migrateurs p?lagiques (thons,
espadons) ? Le retour au r?cif est-il une adaptation globale de chaque population
? son environnement hydrodynamique, comme le sugg?re la th?orie de Michael
Sinclair, de l'universit? de l'?tat de Washington(11)? Nul ne le sait pour
l'instant car la m?thodologie des ?tudes est extr?mement complexe, compte
tenu de la multiplicit? des param?tres naturels mis en jeu. Par exemple,
il n'est pas encore possible de suivre les larves de fa?on continue depuis
l'oc?an jusqu'aux r?cifs. Nos recherches ont malgr? tout permis de comprendre
comment certaines esp?ces colonisent la zone de contact entre l'oc?an et
le r?cif, l? o? d?ferlent les vagues. En fixant des filets dans cette
zone tr?s agit?e, nous avons montr? que les larves pr?tes ? se m?tamorphoser
p?n?trent dans le r?cif la nuit(12). Ce ph?nom?ne prend parfois des allures
spectaculaires - un nombre consid?rable de larves franchissent soudain cette
cr?te et il devient possible de les observer sous l'eau avec un masque et
une torche, lorsque l'oc?an est calme. L' horaire est pr?cis ; les premi?res
larves arrivent dix minutes apr?s le coucher du soleil, avant que les poissons
diurnes du r?cif aient cess? leur activit?. La surface de l'eau cr?pite
alors des multiples attaques des pr?dateurs sur les jeunes recrues. Ensuite,
lorsque la nuit est tomb?e et que les poissons diurnes ont stopp? toute
activit?, la colonisation continue avec moins de risques. Tant qu'il n'y
a pas de lune, elle est intensive; d?s que la lune se l?ve, elle diminue
fortement. La colonisation du r?cif par les larves de poissons est donc r?gul?e
par la lumi?re ambiante. Dans la j ourn?e ou lors de la pleine lune, elle
est quasi inexistante; s'il fait noir sur le r?cif, les larves y arrivent
en grand nombre. Comme dans le cas des tourbillons situ?s en aval des r?cifs,
ce m?canisme va ?galement dans le sens de la th?orie de Johannes : la colonisation
du r?cif a lieu lorsque les risques de pr?dation diminuent. Enfin, nous
avons constat? que la m?tamorphose se produit souvent d?s que les poissons
sont au contact du r?cif. Cela confirme que des stimuli provenant de l'environnement
du r?cif sont impliqu?s dans la m?tamorphose des larves. Des mod?les math?matiques
fond?s sur le recrutement et sur les principaux caract?res biologiques des
esp?ces (courbe de croissance, courbe de survie, etc.) permettent depuis
quelques ann?es de pr?dire l'?volution des stocks de poissons et de mieux
organiser l'effort de p?che. Malgr? tout, un certain nombre de p?cheries
se sont effondr?es ces derni?res ann?es. Il existe bien entendu de multiples
facteurs, techniques, ?conomiques et sociaux pour expliquer ces d?sastres,
souvent li?s ? la surexploitation d'un grand nombre de stocks de poissons;
mais l'effondrement de certains stocks a en fait aussi ?t? observ? alors
que l'effort de p?che n'avait pas ?t? intensif. Les ?v?nements qui influencent
la survie des poissons durant la phase larvaire lors du recrutement (pr?dation,
nourriture, conditions climatiques) apparaissent maintenant comme l'explicatif
ultime de ces oscillations irr?guli?res. Jusque dans les ann?es 1975, les
populations des poissons coralliens ?tait consid?r?es comme un mod?le
de stabilit? du fait des conditions constantes suppos?es du r?cif. Le recrutement
ne semblait pas influencer l'abondance des poissons. Des ?tudes plus pr?cises
sur ces peuplements ont alors fait appara?tre qu'il pouvait varier de fa?on
importante et impr?dictible. L'hypoth?se d'un ?quilibre ?cologique d?termin?
par les fluctuations du recrutement a ?t? ?nonc? en 1980 par Peter Sale,
? l'Universit? du New Hampshire. Le r?le dans cet ?quilibre des ?v?nements
survenant dans le r?cif (comme pr?dation exerc?e sur les juv?niles et
les adultes) par rapport ? ceux ayant lieu dans l'oc?an (comme la mortalit?
larvaire) restait n?anmoins difficile ? appr?hender. En 1994, l'un d'entre
nous (Peter Doherty) et l'un de ses coll?gue Tony Fowler, ont r?alis? la
premi?re d?monstration ? grande ?chelle de l' influence du recrutement
sur l'abondance des populations de poissons dans r?cifs (13). Ils ont ?tudi?
ann?e apr?s ann?e le recrutement des larves du poisson demoiselle Pomacentrus
moluccensis, parall?lement ? l'abondance des adultes dans neuf r?cifs
de la Grande Barri?re d'Australie. Ils ont d?termin? la courbe d?mographique
des populations install?es (en comptant les minuscules stries journali?res
d'accroissement qui se dessinent sur des pi?ces calcifi?es, les otolithes,
situ?es dans l'oreille interne des larves). R?sultat : les fluctuations
du nombre de recrues dans l'espace et dans le temps influencent durablement
ces populations (via la pyramide des ?ges), et la mortalit? naturelle ne
compense pas ces oscillations. L'abondance des poissons dans un r?cif ne
d?pend donc pas seulement de la comp?tition entre les adultes, mais ?galement
du taux de recrutement. Bien des aspects biologiques et ?cologiques de la
vie des poissons des r?cifs coralliens d?pendent ? la fois des ?v?nements
qui ont lieu durant la phase larvaire et de leur passage ? la vie dans les
r?cifs, qui conditionne la quantit? et la diversit? de poissons. L'exploitation
raisonn?e des poissons coralliens ? des fins alimentaires exige de contr?ler
le taux de mortalit? des larves au moment de leur m?tamorphose et des premi?res
heures de la vie dans le r?cif. En m?me temps, comme la densit? naturelle
des poissons est souvent en de?? de la capacit? de charge du r?cif, il
est envisageable d'augmenter artificiellement le taux de recrutement dans
des sites appropri?s. L'?tude de la colonisation du r?cif par les larves
a en effet montr? qu'elles sont plus nombreuses que les juv?niles install?s
quelques jours plus tard. La mortalit? des juv?niles semble extr?mement
importante imm?diatement apr?s la colonisation du r?cif et diminuerait
ensuite de fa?on exponentielle.
Les lagons pourraient servir ? ?laborer une aquaculture extensive
? partir des larves de poissons venant coloniser le r?cif.
|
POUR EN SAVOIR PLUS C. P?tron et C. Gabri?, R?cifs, le monde du corail, Deno?l, 1990.
|
Il para?t donc possible de collecter les larves vivantes d?s leur arriv?e sur le r?cif, de les laisser ensuite se d?velopper dans des sites particuliers sans pr?dateur, afin d'augmenter certains stocks dans des structures isol?es ou ferm?es, ou pour concentrer l'effort de p?che dans des zones adapt?es. Par ailleurs, les lagons des r?cifs coralliens ?tant des milieux relativement clos, ceux-ci pourraient servir ? ?laborer une aquaculture extensive ? partir des larves de poissons collect?es lors de la colonisation. Et le public de plus en plus nombreux sensible ? la magnificence des lagons coralliens trouverait l? un centre d'int?r?t non n?gligeable. L'?tude des poissons des r?cifs coralliens s'av?re non seulement utile pour comprendre des ?cosyst?mes d'une incomparable richesse, mais aussi pour ?lucider certaines lois r?gissant la vie des animaux marins. La diversit?, l'abondance et l'accessibilit? des peuplements de poissons r?cifaux en font d'excellents outils exp?rimentaux pour l'?tude de l'?cologie et de la biologie marine, et des candidats de choix pour une meilleure gestion et une exploitation plus rationnelle des ressources marines vivantes.
Vincent DUFOUR, Serge PLANES et Peter DOHERTY
R?F?RENCES
(1) P.F Sale (ed.) The ecology of fishes on coral reefs Academic Press, New
York, 1991.
(2) M.L. Harmelin-Vivien et F. Bourli?re, Vertebrates in complex tropical
systems, Springer-Verlag Berlin, 1989.
(3) R.E. Tresher, Reproduction in reef fishes, T.F.H. Publications Inc., Neptune
City, New Jersey, 1984.
(4) P.J. Doherty et ai., Env. Biot. Fish, 12, 81, 1985.
(5) R.S. Schelterna, Nature, 217,1159, 1968.
(6) G.W. Barlow, Env. Biot. Fish, 6,65,1981.
(7) R.E. Johannes, Env. Biot. Fish, 3,741, 1978.
(8) S. Planes, Marine Ecology Progress Series, 98, 237, 1993.
(9) S. Planes et al., Marine Bioiogy, 117, 665, 1993.
(10) P.J. Doherty et ai., Ecoiogy, 1995, sous presse.
(11) M. Sinclair, Marine Populations, Washington Sea Grant, University of
Washington Press, 1988.
(12) V. Dufour et R. Galzin, Marine Ecology Progress Series, 102, 143, 1993.
(13) P.J. Doherty et T. Fowler, Science, 263, 935, 1994.
(14) G.W. Boelhert et ai., Deep-Sea Research,39, 439,1992.
(15) K. P. Black et aL, Marine Ecology Progress Series, 74,1991